
| Neurociências > Um novo referencial para compreender a gagueira: o modelo pré-motor duplo | |
Per A. ALM
Departamento de Psicologia Experimental, Universidade de Oxford, Reino Unido
ResumoUm modelo explicativo é proposto aqui: a teoria da gagueira baseada nos sistemas pré-motores duplos. Pistas para a temporalização dos segmentos da fala podem ser fornecidas por um de dois sistemas pré-motores paralelos, o lateral e o medial. A gagueira parece estar relacionada a pistas defeituosas produzidas pelo sistema medial (que inclui os núcleos da base). Um importante fator para isso pode ser um input deficiente produzido pelo córtex motor em direção aos núcleos da base, deixando-os fora de controle e sujeitos a influências emocionais desproporcionalmente fortes do sistema límbico. A maioria das condições indutoras de fluência caracterizam-se pela mudança do centro de controle de temporalização da fala do sistema medial para o sistema lateral. 1. IntroduçãoA gagueira é uma desordem motora da fala insuficientemente compreendida. Há uma série de observações e descobertas experimentais sobre gagueira que precisam ser conectadas por um novo referencial explicativo que consiga unir as peças espalhadas num todo coerente. Um dos principais aspectos da gagueira são as várias condições que tendem a aliviá-la temporariamente: o efeito rítmico, a leitura em coro, o canto, a imitação de um sotaque estrangeiro, o feedback auditivo alterado, etc. O efeito freqüentemente notável dessas condições fornece indícios importantes da provável natureza básica da gagueira. Outras peças importantes do quebra-cabeça são as informações provenientes da neuroimagem, o efeito de agentes farmacológicos, os sintomas exibidos no momento da gagueira, a localização de lesões que causam gagueira em adultos, o padrão típico de surgimento e remissão da gagueira na infância e a influência de fatores emocionais relatada com freqüência por pessoas que gaguejam. O objetivo desta apresentação é propor um novo modelo explicativo, denominado teoria da gagueira baseada nos sistemas pré-motores duplos. O modelo é primariamente baseado no raciocínio teórico contido em Alm (2004), que considera o papel dos núcleos da base na gagueira, e nas análises adicionais contidas em Alm (2005). Neste último trabalho, o papel dos núcleos da base na gagueira foi colocado no referencial mais amplo fornecido pela hipótese dos sistemas pré-motores duplos, formulada por Goldberg (1985, 1990) e Passingham (1987). 2. Núcleos da base e gagueiraHistórico. A idéia de que a gagueira está relacionada a distúrbios no funcionamento dos núcleos da base não é nova. Já por volta de 1920 o médico alemão Sahli fez esta proposição, baseado em observações da encefalite epidêmica. Na literatura mais contemporânea sobre gagueira, os núcleos da base apareceram em Rosenberger (1980), na discussão do efeito de bloqueadores do receptor de dopamina para aliviar a gagueira. A dopamina é o principal regulador das funções dos núcleos da base. Em 1987, Ludlow et al. publicaram um estudo detalhado sobre a localização de lesões que provocaram gagueira em adultos. As únicas estruturas da substância cinzenta que estavam, com maior freqüência, significativamente afetadas no grupo com gagueira eram partes dos núcleos da base. Wu et al. (1995) relataram metabolismo reduzido em uma parte do lado esquerdo dos núcleos da base e, mais tarde, o mesmo grupo encontrou uma captação extremamente elevada do precursor de dopamina em um pequeno estudo de três adultos com gagueira (Wu et al., 1997). Molt (1999) discutiu possíveis ligações entre gagueira e núcleos da base, baseado nas semelhanças entre gagueira e síndrome de Tourette, distonia e outros distúrbios neurológicos do movimento. Vários autores têm feito correlação entre distonias ocupacionais e gagueira (Bluemel, 1930; Kiziltan & Akalin, 1996; Victor & Ropper, 2001). Anatomia e pistas de temporalização. Em Alm (2004), o possível papel dos núcleos da base na gagueira foi revisado em detalhes e implicações teóricas foram propostas. Em resumo, os núcleos da base são estruturas no centro do cérebro envolvidas na automatização de uma grande quantidade de funções, desde funções cognitivas e motivacionais, até o controle motor. Eles recebem input da maioria das partes do córtex cerebral e do sistema límbico e projetam-se principalmente para o córtex frontal (veja figura 1). Desta forma, os núcleos da base modulam o estado das regiões do córtex frontal. Especialmente interessante para a discussão sobre gagueira é o fato de os núcleos da base terem um papel chave na automatização de seqüências motoras rápidas. A fala é uma seqüência motora em que os submovimentos precisam de sincronização exata, ou seja, sinais precisos de temporalização e disparo para o surgimento dos elementos da fala. Há forte indicação de que a área motora suplementar (AMS) desempenha um papel fundamental nesta sincronização dos elementos motores da fala e que os núcleos da base normalmente auxiliam esse processo fornecendo pistas de temporalização para a AMS.
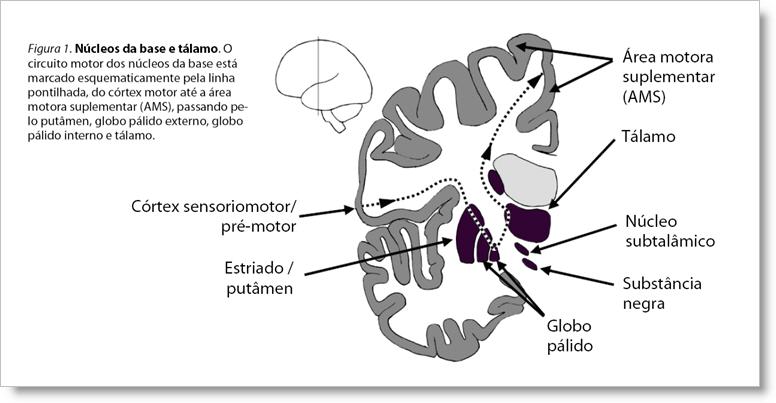 Ativação focal e inibição difusa do córtex. Os circuitos dos núcleos da base são organizados em uma via direta e uma via indireta. Essas duas vias trabalham em sinergia para modular a atividade do córtex frontal: a via indireta fornece uma inibição difusa da atividade cortical e a via direta, uma ativação focal da ação desejada (Mink & Thach, 1993). Além disso, as duas vias são dominadas por tipos distintos de receptores de dopamina, D1 vs. D2, resultando em efeitos diferenciados do neurotransmissor. O fornecimento de pistas de temporalização para a AMS é uma função dos núcleos da base que depende de uma distinção clara entre ativação focal e inibição difusa do córtex; em outras palavras, uma boa razão sinal-ruído. Com base neste modelo, fica fácil imaginar que as pistas de temporalização dos núcleos da base podem ser distorcidas de diferentes formas. Uma ativação focal muito fraca da via direta resultaria numa ativação deficiente da ação desejada, levando, por exemplo, a dificuldades na iniciação dos movimentos da fala. Por outro lado, problemas na inibição difusa do córtex, fornecida pela via indireta, poderia resultar numa situação que combinaria liberação de movimentos involuntários e dificuldade de liberação de movimentos voluntários. Esses cenários têm paralelos evidentes com os sintomas da gagueira. Os distúrbios motores dos núcleos da base são caracterizados por problemas na iniciação motora, movimentos involuntários e tensão muscular desregulada, freqüentemente com contração concomitante de músculos antagonistas. Influências emocionais sobre os sintomas. Outra característica dos distúrbios motores dos núcleos da base é a influência emocional sobre os sintomas, com a tensão nervosa tornando-os piores e o relaxamento induzindo alguma melhora (Victor and Ropper, 2001). Essa também é uma experiência clínica comum da gagueira, freqüentemente mencionada por quem gagueja. Efeitos de drogas. As drogas com o mais forte efeito sobre a gagueira, para melhor ou para pior, tipicamente afetam o neurotransmissor dopamina (Alm, 2004). A dopamina tem efeitos em muitas partes do cérebro, mas os efeitos mais explícitos estão nos núcleos da base, com influência imediata e freqüentemente dramática nos movimentos, no humor e no nível geral de atividade. Os efeitos da dopamina sobre a gagueira parecem ser complexos e heterogêneos. Tanto a inibição quanto a ativação da dopamina podem fazer a gagueira piorar ou melhorar em diferentes casos. Início precoce e remissão. A gagueira freqüentemente inicia por volta dos 2,5 e 3 anos de idade, com um elevado percentual de remissões espontâneas ainda nessa fase. O único processo de desenvolvimento neurológico que parece se ajustar a esse padrão, com um pico aos 2,5-3 anos de idade, é o número de receptores de dopamina nos núcleos da base (veja fig. 2 em Alm, 2004). De acordo com o modelo dos núcleos da base discutido acima, um elevado número de receptores D2 resultará numa redução da inibição geral do córtex. Além disso, uma baixa razão entre receptores D1/D2 levaria a um prejuízo da relação sinal-ruído no output dos núcleos da base. Os dados disponíveis, ainda muito limitados, sugerem que a relação D1/D2 tende a ser mais baixa em meninos do que em meninas na infância, possivelmente influenciando a prevalência de gênero na gagueira. Para resumir, parece possível que o padrão típico de surgimento e remissão da gagueira na infância esteja relacionado em grande escala a uma fase natural no desenvolvimento do sistema dopaminérgico nos núcleos da base. Condições indutoras de fluência. O último aspecto ligando a gagueira ao sistema dos núcleos da base são as condições indutoras de fluência, como o efeito do ritmo (falar seguindo um ritmo externo). O mal de Parkinson é causado por níveis insuficientes de dopamina nos núcleos da base, resultando em problemas de iniciação motora e tensão muscular desregulada. É interessante notar que o efeito do ritmo é também uma característica do mal de Parkinson. Em parkinsonianos também ocorre um prejuízo na capacidade de realizar movimentos auto-iniciados, mas a performance é sensivelmente melhorada se os movimentos são guiados por estímulos rítmicos externos. Este padrão de dificuldade nos movimentos auto-iniciados também foi observado em macacos depois de lesões experimentais no putâmen (uma parte dos circuitos motores dos núcleos da base) ou na AMS (Nixon & Passingham, 1998; Thaler, Chen, Nixon, Stern, & Passingham, 1995). Por outro lado, lesões do córtex pré-motor lateral não resultaram neste tipo de dificuldade (Thaler et al., 1995). Em resumo, o efeito do ritmo sobre a gagueira fornece uma forte indicação de que a dificuldade está relacionada às funções do sistema motor que envolve a participação dos núcleos da base e da área motora suplementar (AMS). O efeito do feedback auditivo alterado (FAA) sobre a gagueira ainda é intrigante e mal compreendido. Pistas para a elucidação do mecanismo envolvido podem ser fornecidas por estudos que relatam melhora na fala de pacientes com mal de Parkinson e dificuldade de fala associada. Essas melhoras foram obtidas por feedback auditivo atrasado (DAF) e por feedback auditivo com freqüência alterada (FAF) (Downie, Low, & Lindsay, 1981; Brendel, Lowit, & Howell, 2004). 3. O modelo dos sistemas pré-motores duplosA hipótese dos sistemas pré-motores duplos. Em Alm (2005), o modelo explicativo da gagueira relacionado a anormalidades no funcionamento dos núcleos da base foi desenvolvido ainda mais e o papel dos núcleos da base foi posto no referencial mais amplo fornecido pela hipótese dos sistemas pré-motores duplos formulada por Goldberg (1985, 1990) e Passingham (1987). Este modelo enfatiza que as funções motoras dos núcleos da base integram um sistema maior, o sistema pré-motor medial, que opera em paralelo com o sistema pré-motor lateral. Os principais componentes do sistema medial são os núcleos da base e a AMS, enquanto o sistema lateral compõe-se do córtex pré-motor lateral e do cerebelo. (v. fig. 2). Esses sistemas parecem operar sob diferentes condições, com o sistema medial produzindo ações auto-iniciadas baseadas em um modelo interno do mundo, numa relação íntima com o sistema límbico e com fatores motivacionais. Por outro lado, o sistema lateral tipicamente opera em relação ao input sensorial e se baseia no controle de feedback, havendo um controle voluntário e consciente da ação. A distinção entre esse dois sistemas também foi reconhecida por outros autores. Damasio (1994), por exemplo, afirmou que um sorriso verdadeiro é produzido pela atividade de estruturas no sistema medial, enquanto uma tentativa não-emocional de sorrir para um fotógrafo é controlada por estruturas do sistema lateral.
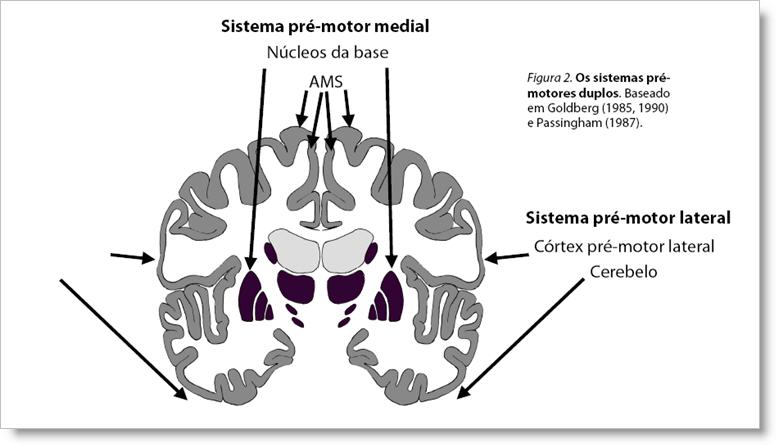 Os sistemas duplos no processamento da fala. Goldberg (1985) discutiu o papel desses dois sistemas na fala, mas não chegou a relacionar diretamente seu modelo à gagueira. De acordo com Goldberg, os dois sistemas são capazes de controlar a temporalização dos segmentos motores da fala, mas sob condições diferentes. Na fala proposicional espontânea, o controle da temporalização é normalmente direcionado para o sistema medial. Por outro lado, quando a temporalização dos segmentos da fala está ligada a estímulos externos (como um metrônomo), o processo é executado pelo sistema lateral. Isto explicaria o efeito do ritmo sobre a gagueira e o mal de Parkinson, e também o efeito da fala em coro, no qual se baseiam os aparelhos eletrônicos intrauriculares indutores de fluência: a dificuldade localizada no sistema medial seria, sob a influência desses estímulos, contornada pela mudança do controle da fala para o sistema lateral. Imitação, encenação e canto. O sistema lateral também tem a capacidade de controlar a temporalização da fala sem input externo explícito, mas isso requer um nível aumentado de atenção consciente para alguns aspectos da fala, no sentido de torná-la menos automática. Este efeito pode ser demonstrado pela redução da gagueira quando se imita um sotaque estrangeiro, quando se encena um personagem no teatro, quando se fala de forma exageradamente ritmada ou quando se utiliza uma taxa de elocução controlada conscientemente. Além disso, foi mostrado que cantar ou falar de forma exageradamente ritmada são atividades que estão relacionadas a uma atividade aumentada no córtex pré-motor lateral direito (Riecker et al., 2000; Riecker et al., 2002), indicando de fato uma mudança do centro de controle da fala para o sistema pré-motor lateral. Parece provável que esteja aí a explicação para o fato amplamente conhecido de a gagueira estar ausente quando se canta: a temporalização das sílabas na canção é executada por um sistema que normalmente não está prejudicado em pessoas com gagueira. Feedback auditivo alterado. Foi demonstrado que o feedback auditivo alterado (por atraso ou mudança de freqüência, DAF e FAF, respectivamente) provoca uma atividade aumentada no córtex auditivo e no cerebelo, tanto para pessoas que gaguejam quanto para as do grupo controle (Watkins, Davis, & Howell, 2005). Em relação ao modelo dos sistemas pré-motores duplos, esta descoberta indica que o feedback auditivo alterado ativa o sistema lateral, aumentado o nível de processamento do input auditivo. Isso poderia explicar o efeito de facilitação que o feedback auditivo alterado causa tanto em pacientes com gagueira quanto em pacientes com mal de Parkinson: o mau funcionamento do sistema medial seria contornado pela mudança do controle da fala para o sistema lateral. 4. Problemas no input motor dos núcleos da base?O modelo dos sistemas pré-motores duplos enfatiza que os núcleos da base são parte de um sistema maior, o sistema pré-motor medial. Isto significa que problemas na função motora dos núcleos da base podem ser resultado de perturbações em diferentes partes deste circuito e isso inclui problemas no input do córtex em direção aos núcleos da base, problemas nos próprios núcleos da base, problemas no tálamo, na AMS e em todos os tratos fibrosos de substância branca conectando essas estruturas. Anormalidades estruturais causando input motor deficiente nos núcleos da base? Alguns estudos recentes indicam a existência de anomalias estruturais em uma região cortical envolvendo a área sensório-motora esquerda relacionada à face e à laringe e também o córtex pré-motor esquerdo ventral, além de anomalias na substância branca logo abaixo dessas regiões corticais, veja figura 3. (Foundas et al., 2001; Sommer et al., 2002; Jancke et al., 2004; Watkins et al., 2006).
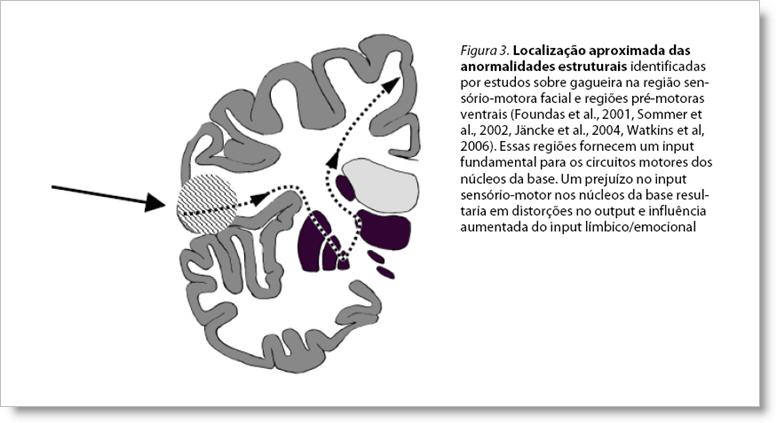 Contudo, não está claro como essas descobertas se relacionam com o cenário mais amplo da gagueira: os sintomas, os efeitos das drogas dopaminérgicas, as condições indutoras de fluência, etc. Uma interpretação dessas descobertas foi sugerida em Alm (2004): os núcleos da base dependem do input das regiões motoras do lado esquerdo para produzir pistas de temporalização para a iniciação do próximo segmento motor. Se o input que os núcleos da base recebem a partir das regiões motoras for desorganizado, a função dos núcleos da base ficará fora de controle, possivelmente resultando em gagueira. Input motor reduzido aumentando a influência emocional sobre os núcleos da base. Os circuitos motores dos núcleos da base recebem projeções do sistema límbico, que fornece inputs relacionados ao estado emocional. Se o input motor do córtex para os núcleos da base estiver reduzido, é provável que o input fornecido pelo sistema límbico adquira uma influência desproporcionalmente forte sobre o sistema motor da fala nos núcleos da base, tornando a fluência mais vulnerável aos influxos emocionais. Este modelo prevê que o grau de influência emocional sobre os sintomas da gagueira dependerá da localização do distúrbio no sistema pré-motor medial. Se a gagueira for causada por uma disfunção situada após os núcleos da base na alça que compõe o sistema medial (ou seja, no tálamo ou na AMS), os sinais emocionais que chegam aos núcleos da base também serão afetados e os fatores emocionais não terão forte influência sobre os sintomas. 5. Pesquisa futura: um possível conflito inter-hemisférico dos núcleos da base?Em Alm (2004), argumentei contra a hipótese de a gagueira estar relacionada a um conflito pelo controle da fala entre os hemisférios cerebrais esquerdo e direito. Ao contrário, defendi que um envolvimento maior do hemisfério direito na gagueira estivesse compensando a dificuldade localizada no hemisfério esquerdo. Contudo, recentemente comecei a considerar a possibilidade de que outputs conflitantes provenientes dos núcleos da base nos hemisférios esquerdo e direito em direção à AMS poderiam ter um papel importante na gagueira, especialmente para as posturas fixas (bloqueios e prolongamentos). Parece provável que o sistema dos núcleos da base no hemisfério esquerdo tenha normalmente um papel dominante no seqüenciamento motor da fala. Se o sistema no lado direito tenta compensar uma instabilidade no sistema do lado esquerdo, o resultado pode ser um output bilateral assíncrono dos núcleos da base para a AMS, possivelmente causando uma ruptura do seqüenciamento motor da fala. Argumentos para esta hipótese serão apresentados futuramente. 6. ResumoA teoria aqui proposta para a gagueira, a teoria dos sistemas pré-motores duplos, é uma expansão da teoria anterior, que relacionava a gagueira a uma disfunção dos núcleos da base. Essa expansão enfatiza que as funções motoras dos núcleos da base são dependentes de um sistema maior, o sistema pré-motor medial, incluindo o input a partir de regiões do córtex motor lateral. A teoria do sistema pré-motor duplo explica a maioria das condições indutoras de fluência como resultado de uma mudança no controle de temporalização da fala do sistema medial para o sistema lateral, o que contornaria a suposta instabilidade presente no sistema medial. A gagueira ocorreria como resultado de vários fatores afetando o sistema pré-motor medial, por exemplo: deficiência no input originário das regiões do córtex motor para os núcleos da base, baixa relação entre receptores D1/D2 no estriado, lesões focais do estriado ou outras partes do sistema medial, etc. Também se especula a possibilidade de um output bilateral conflitante dos núcleos da base ter um papel na gagueira. ReferênciasAlm, P. A. (2004). Stuttering and the basal ganglia circuits. Journal of Communication Disorders, 37, 325-369. Alm, P. A. (2005). On the causal mechanisms of stuttering. Doctoral dissertation, Dept. of Clinical Neuroscience, Lund University, Sweden. Bluemel, C. S. (1930). Mental aspects of stammering. Baltimore, Williams and Wilkins. Brendel, B., Lowit, A., & Howell, P. (2004). The effects of delayed and frequency shifted feedback on speakers with Parkinsons disease. Journal of Medical Speech Language Pathology, 12, 131-138. Damasio, A. R. (1994). Descartes error: emotion, reason, and the human brain. N.Y.: Grosset/Putnam. Downie, A. W., Low, J. M., & Lindsay, D. D. (1981). Speech disorder in parkinsonism - usefulness of delayed auditory feedback in selected cases. British Journal of Disorders of Communication, 16, 135-139. Foundas, A. L., Bollich, A. M., Corey, D. M., Hurley, M., & Heilman, K. M. (2001). Anomalous anatomy of speech-language areas in adults with persistent developmental stuttering. Neurology, 57, 207-215. Goldberg, G. (1985). Supplementary motor area structure and function: Review and hypotheses. The Behavioral and Brain Sciences, 8, 567-616. Goldberg, G., & Bloom K. K. (1990). The alien hand sign. Localization, lateralization and recovery. American Journal of Physical Medicine & Rehabilitation. 69, 228-238. Jancke, L., Hanggi, J., & Steinmetz, H. (2004). Morphological brain differences between adult stutterers and non-stutterers. BMC Neurology, 4, 23. Kiziltan, G. & Akalin, M. A. (1996). Stuttering may be a type of action dystonia. Movement Disorders, 11, 278-282. Ludlow, C. L., Rosenberg, J., Salazar, A., Grafman, J., & Smutok, M. (1987). Site of penetrating brain lesions causing chronic acquired stuttering. Annals of Neurology, 22, 60-66. Mink, J. W. & Thach, W. T. (1993). Basal ganglia intrinsic circuits and their role in behavior. Current Opinion in Neurobiology, 3, 950-957. Molt LF (1999). The basal ganglias possible role in stuttering. Proc. 2nd Internat. Stuttering Awareness Day, Internet. www.mnsu.edu/comdis/isad2/papers/molt2.html Nixon, P. D. & Passingham, R. E. (1998). The striatum and self-paced movements. Behavioral Neuroscience, 112, 719-724. Passingham, R. E. (1987). Two cortical systems for directing movement. Ciba Foundation Symposium, 132, 151-164. Riecker, A., Ackermann, H., Wildgruber, D., Dogil, G., & Grodd, W. (2000). Opposite hemispheric lateralization effects during speaking and singing at motor cortex, insula and cerebellum. Neuroreport, 11, 1997-2000. Riecker, A., Wildgruber, D., Dogil, G., Grodd, W., & Ackermann, H. (2002). Hemispheric lateralization effects of rhythm implementation during syllable repetitions: an fMRI study. Neuroimage, 16, 169-176. Rosenberger, P. B. (1980). Dopaminergic systems and speech fluency. Journal of Fluency Disorders, 5, 255-267. Sommer, M., Koch, M. A., Paulus W., Weiller, C., & Buchel, C. (2002). Disconnection of speech-relevant brain areas in persistent developmental stuttering. Lancet, 360, 380-383. Thaler, D., Chen, Y. C., Nixon, P. D., Stern, C. E., & Passingham, R. E. (1995). The functions of the medial premotor cortex. I. Simple learned movements. Experimental Brain Research, 102, 445-460. Victor, M. & Ropper, A. H. (2001). Adams and Victors principles of neurology. (7 ed.) N.Y.: McGraw Hill. Watkins K, Davis M, & Howell P (2005). Brain activity during altered auditory feedback. Paper presented at the Oxford Dysfluency Conference 2005, Oxford. Watkins, K., Smith, S., Davis, S., & Howell, P. (2006). Structural and functional brain abnormalities associated with developmental stuttering. Poster presented at Human Brain Mapping, Florence, June 11-15, 2006. Wu, J. C., Maguire, G., Riley, G., Fallon, J., LaCasse, L., Chin, S., Klein, E., Tang, C., Cadwell, S., & Lottenberg, S. (1995). A positron emission tomography [18F]deoxyglucose study of developmental stuttering. Neuroreport, 6, 501-505. Wu, J. C., Maguire, G., Riley, G., Lee, A., Keator, D., Tang, C., Fallon, J., & Najafi, A. (1997). Increased dopamine activity associated with stuttering. Neuroreport, 8, 767-770.
Traduzido por Hugo Silva e revisado por Sandra Merlo. |
|
| Voltar |
|
| "Instituto Brasileiro de Fluência - IBF" Todos os direitos reservados. |